A unit of meaning, with a structure
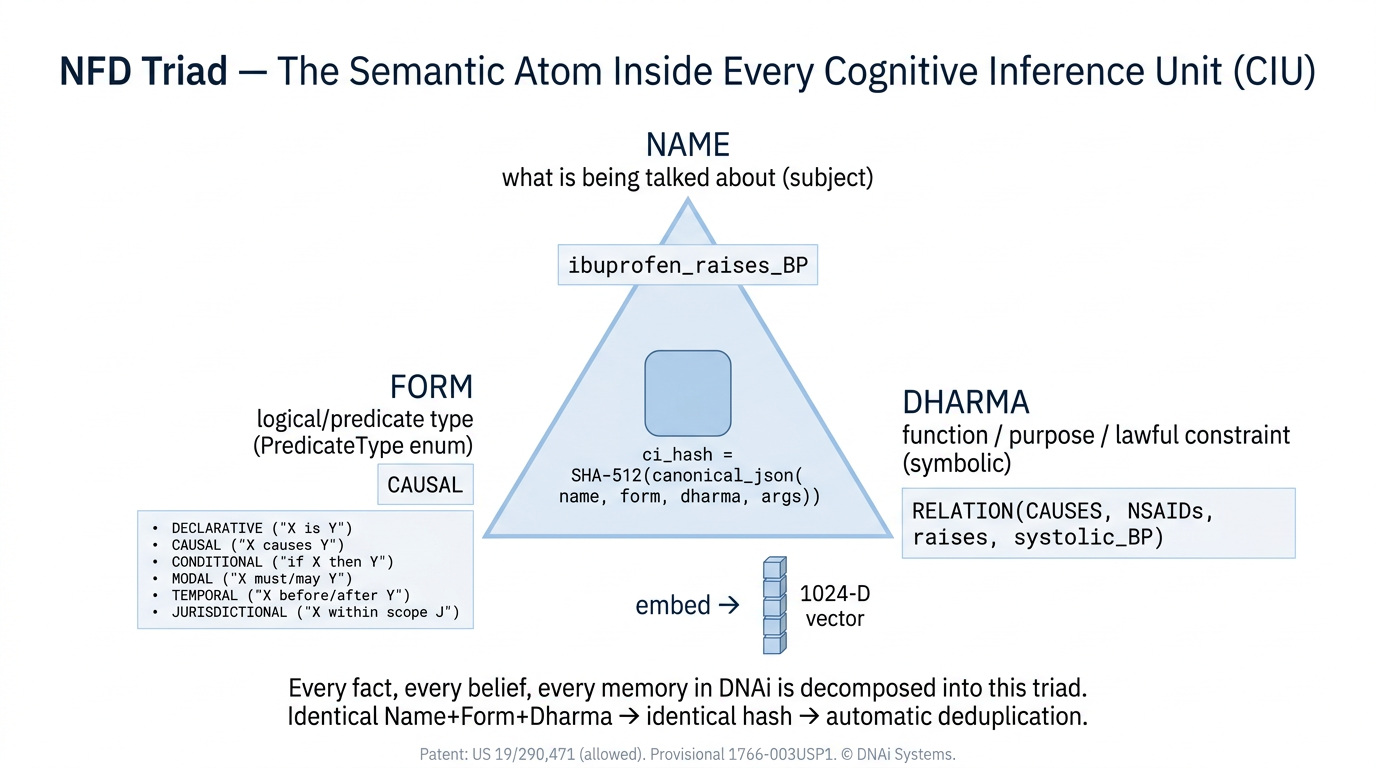
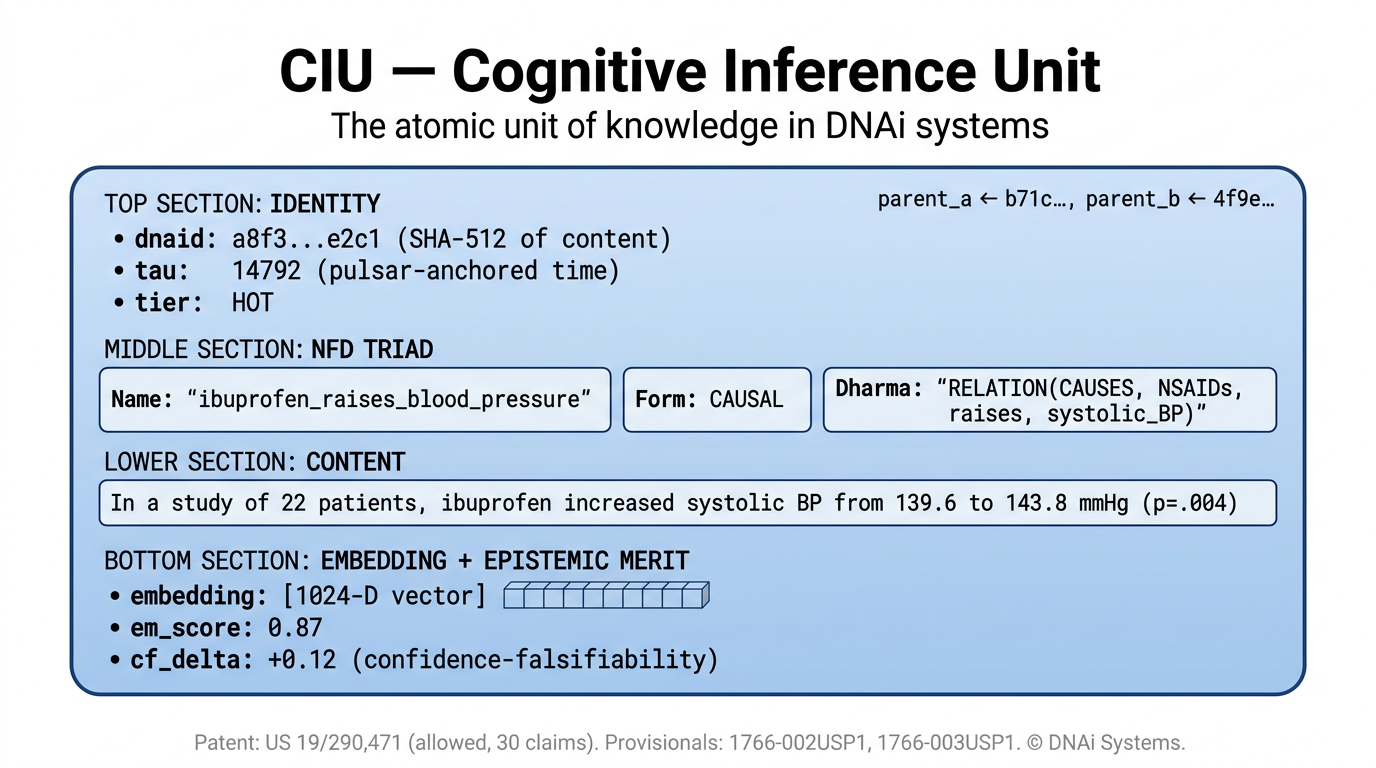
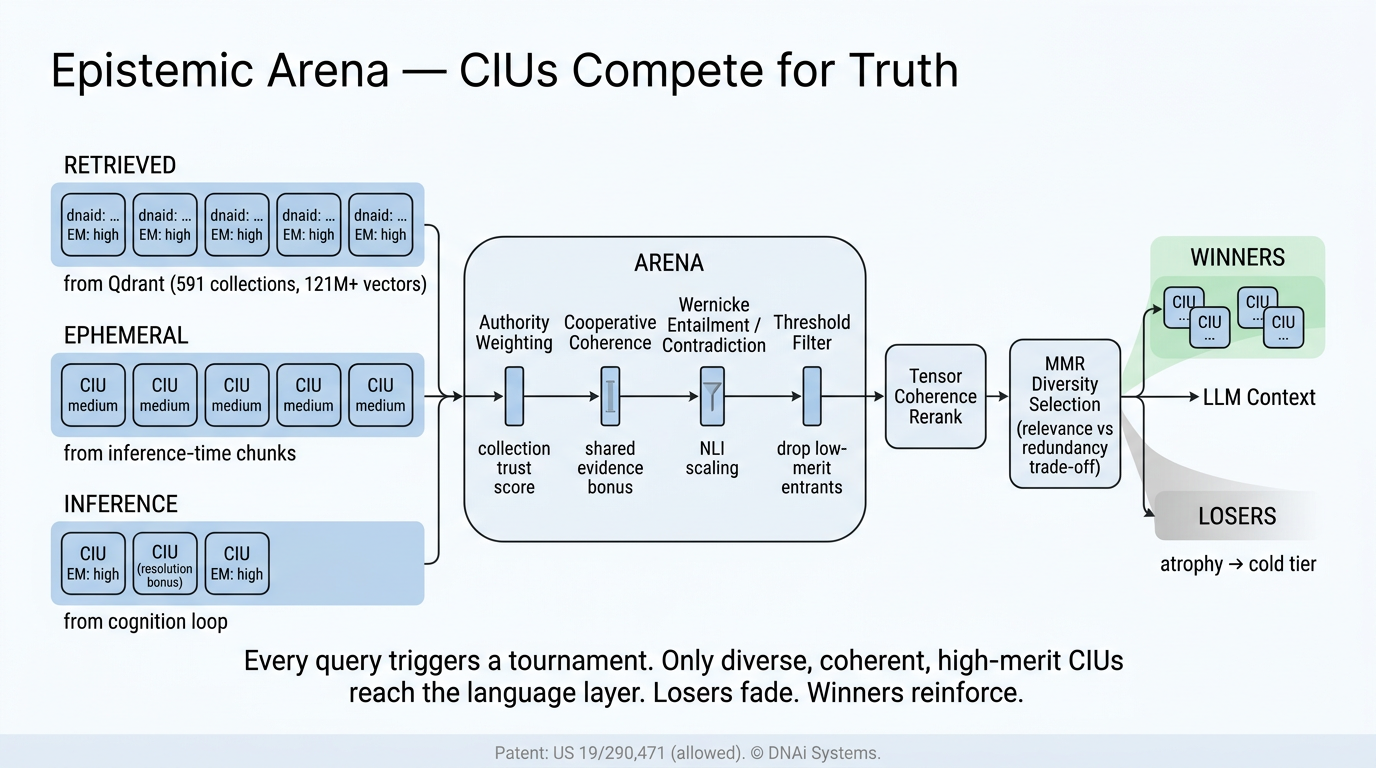
The atomic unit of DNAi knowledge is the Cognitive Inference Unit, abbreviated CIU. The CIU is the term used in DNAi’s allowed United States patent application 19/290,471 and in the runtime ledger code. A CIU is a propositional commitment, not a free text sentence and not a token, built from a three slot structure called the NFD triad: a Name (the subject), a Form (the predicate type, drawn from a fixed set including declarative, causal, conditional, modal, temporal, and jurisdictional), and a Dharma (the function or lawful constraint the proposition asserts).
Two CIUs that share the same Name, Form, and Dharma collapse to the same content addressed identity. Identity is semantic, not just lexical. The runtime canonicalizes Name through entity resolution against the indexed knowledge base (so aspirin, acetylsalicylic acid, and ASA resolve to one Name) before computing the SHA‑512 digest used as identity and as the input to the parent hash chain in the ledger described in Section 3. Each CIU also carries a tier label (HOT, WARM, or COLD), a confidence falsifiability delta (CFΔ), and a contradiction score, and is created, promoted, demoted, or atrophied as a single addressable entity.
Brain biology offers a structural analogue at the population level rather than the single neuron level. The CIU is the addressable digital counterpart, with deterministic identity via cryptographic hash rather than statistical immediate early gene staining.